Dynamic species distribution models of Antarctic blue whales in the Weddell Sea using visual sighting and passive acoustic monitoring data
Aim: Species distribution models (SDMs) are essential tools in ecology and conservation. However, the scarcity of visual sightings of marine mammals in remote polar areas hinders the effective application of SDMs there. Passive acoustic monitoring (PAM) data provide year-round information and overcome foul weather limitations faced by visual surveys. However, the use of PAM data in SDMs has been sparse so far. Here, we use PAM-based SDMs to investigate the spatiotemporal distribution of the critically endangered Antarctic blue whale in the Weddell Sea.
Location: The Weddell Sea
Methods: We used presence-only dynamic SDMs employing visual sightings and PAM detections in independent models. We compared the two independent models with a third combined model that integrated both visual and PAM data, aiming at leveraging the advantages of each data type: the extensive spatial extent of visual data and the broader temporal/environmental range of PAM data.
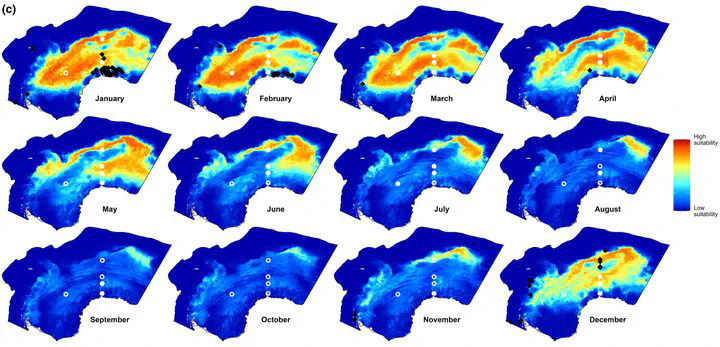
Results: Visual and PAM data prove complementary, as indicated by a low spatial overlap between daily predictions and the low predictability of each model at detections of other data types. Combined data models reproduced suitable habitats as given by both independent models. Visual data models indicate areas close to the sea ice edge (SIE) and with low-to-moderate sea ice concentrations (SIC) as suitable, while PAM data models identified suitable habitats at a broader range of distances to SIE and relatively higher SIC.
Main conclusions: The results demonstrate the potential of PAM data to predict year-round marine mammal habitat suitability at large spatial scales. We provide reasons for discrepancies between SDMs based on either data type and give methodological recommendations on using PAM data in SDMs. Combining visual and PAM data in future SDMs is promising for studying vocalized animals, particularly when using recent advances in integrated distribution modeling methods.