Multi-year presence of humpback whales in the Atlantic sector of the Southern Ocean but not during El Niño
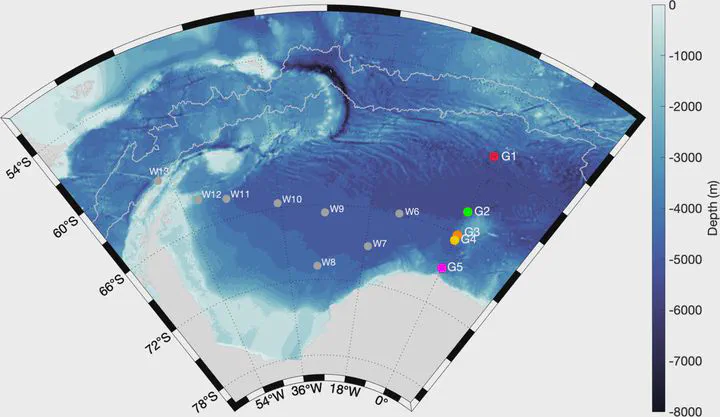
Humpback whales are thought to undertake annual migrations between their low latitude breeding grounds and high latitude feeding grounds. However, under specific conditions, humpback whales sometimes change their migratory destination or skip migration overall. Here we document the surprising persistent presence of humpback whales in the Atlantic sector of the Southern Ocean during five years (2011, 2012, 2013, 2017, and 2018) using passive acoustic data. However, in the El Niño years 2015 and 2016, humpback whales were virtually absent. Our data show that humpback whales are systematically present in the Atlantic sector of the Southern Ocean and suggest that these whales are particularly sensitive to climate oscillations which have profound effects on winds, sea ice extent, primary production, and especially krill productivity.