Conserving Egypt's reptiles under climate change
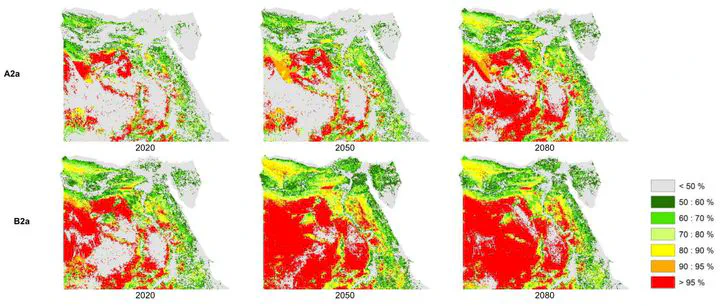
Climate change has caused range shifts and extinctions of many species in the recent past. In this study, the effects of climate change on Egyptian reptiles were investigated for the first time using species distribution models. Maxent was used to model the current and future distributions of suitable habitats for 75 terrestrial reptile species from Egypt. The modelled distribution for current suitable conditions for each species was projected into the future at three time slices using two emission scenarios from four global circulation models and under two assumptions of dispersal ability. Climate change is expected to vary in its effects spatially, with some areas characterized by increased species richness while others show declines. Future range changes vary among species and different future projections, from the entire loss to large gains in range. Two species were expected to become extinct in at least one future projection, and eight species were expected to lose >80% of their current distribution. Although Protected Areas have greater conservation value, on average, compared to unprotected areas, they appear inadequate to conserve Egyptian reptiles under expected climate change.